記憶を担うシナプス受容体が睡眠時に保護されていることを解明
背景 睡眠と記憶の定着
睡眠は様々な認知機能に関与しています。睡眠不足は、反応時間、作業記憶、持続的な注意から、実行制御、感情、気分に至るまで、様々な認知機能を損ないます。一方、睡眠は記憶の定着と統合、要点の抽出、さらに忘却を促し、学習能力を回復させます。これまでに、多数の研究が、長期記憶を担う大脳皮質や海馬において、学習時に活性化した神経細胞が睡眠脳波のリズムで再活性化して記憶の定着を担うことを示しています。特に、私たちは睡眠時の神経活動を操作することによって、記憶の定着に重要な役割を果たす神経活動のタイムコースを解明してきました。その結果、記憶の定着に学習直後の睡眠時における神経活動が重要な役割を果たす一方で、時間経過によって十分に定着した記憶は神経活動の影響を受けにくいことが分かりました。また、神経活動は神経細胞間の情報伝達を行うシナプスの分子構成を再編し、長期シナプス可塑性と呼ばれるシナプスの伝達効率の長期的な変化を生じます。そこで、私たちは、覚醒中の学習時及び学習直後の睡眠時における神経活動がシナプス可塑性1を通じて、持続的な記憶痕跡を形成すると予想しました。
覚醒中の学習は大脳皮質や海馬等の記憶に関連する脳領域の一部のシナプスにおいて、興奮性シナプス伝達を担うAMPA受容体2の量を増加させて、記憶の保持を担うシグナルを形成します。一方、睡眠はAMPA受容体を平均的に減少させ、脳回路をクールダウンさせて恒常性を維持すると考えられています (図1)。しかし、従来の死後脳における解剖学的手法や生化学的手法では、学習や睡眠によるシナプス・分子動態を追跡することは困難でした。そのため、睡眠がAMPA受容体の減少と記憶の定着をどの様に両立させているのかは不明でした。そこで、私たちは生体マウスにおいて、AMPA受容体を二光子顕微鏡3により経時蛍光イメージングし、学習と睡眠によるシナプス可塑性を個々のシナプスにおいて高解像度で追跡しました。
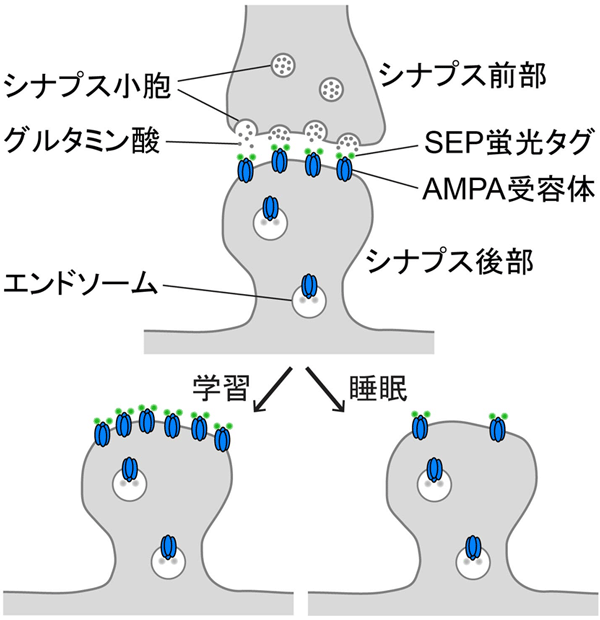 図1. 学習と睡眠によるAMPA受容体量の平均的な変化
図1. 学習と睡眠によるAMPA受容体量の平均的な変化シナプス後部のAMPA受容体量は学習により増加し、睡眠により減少する。本研究は比較的高いpHを有する細胞外液に反応して蛍光を発するSEP蛍光タグを利用して、細胞膜表面の機能的なAMPA受容体を可視化している。
手法 シナプスAMPA受容体の可塑性の生体イメージング法
AMPA受容体のGluA1サブユニットを蛍光可視化するために、SEP (Super Ecliptic pHluorin)-GluA1を用いました。SEPはpH依存型の緑色蛍光タンパク質で、細胞外と細胞内のpHの違いを利用し、細胞膜上の機能的な受容体を選択的に蛍光可視化します。また、神経細胞の形態を蛍光可視化するために、赤色蛍光タンパク質のdsRed2を用いました。SEP-GluA1とdsRed2の神経細胞への発現を誘導するために、14.5日齢の胚に子宮内電気穿孔法4を適用しました。仔マウスの成熟後、脳表にガラス窓を設置しました。そして、二光子顕微鏡を用いて、一次運動皮質の2/3層錐体細胞5の樹状突起スパイン6において、運動学習と睡眠の前後において経時イメージングを行いました (図2)。コンプレックスホイール試験7という運動学習試験を用いて、睡眠が運動記憶を固定化するシナプス・分子メカニズムを調べました。
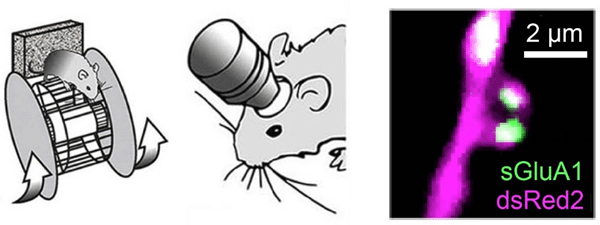 図2. マウスの運動学習とAMPA受容体の生体イメージング
図2. マウスの運動学習とAMPA受容体の生体イメージング運動学習 (左) と睡眠の前後に、二光子顕微鏡 (中) を適用して、個々の樹状突起スパインのAMPA受容体を蛍光可視化した (右)。
結果と考察 学習と睡眠によるシナプス選択的なAMPA受容体の可塑性
予想通り、樹状突起スパインの平均AMPA受容体量は、学習によって増加し、睡眠により減少することを確認しました。運動学習によりAMPA受容体が特に増加した一部のスパインに注目すると、運動学習後の睡眠や断眠の影響を受けずに保護されており、2日目においてもAMPA受容体の増加を持続して生じていました。一方、その他のスパイン (学習によりAMPA受容体の増加しなかったスパイン) のAMPA受容体は運動学習後の睡眠時に減少しました。この運動学習後のAMPA受容体の減少は2日目の運動記憶成績と相関していました。さらに、運動学習後のマウスに断眠処置を行うと、AMPA受容体の減少が生じなくなり、2日目の運動記憶成績が低下しました。これらより、運動学習による一部のスパインにおけるAMPA受容体の増加と、運動学習後の睡眠によるその他のスパインにおけるAMPA受容体の減少との相乗効果が、運動記憶を担うシグナルを相対的に強化していると考えられます (図3)。
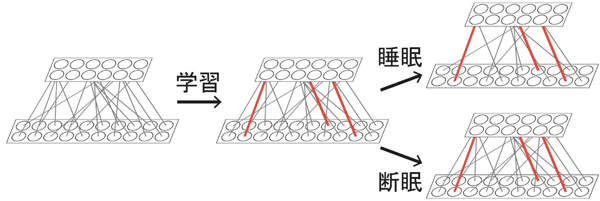 図3. 学習と睡眠による個々のシナプスの可塑性
図3. 学習と睡眠による個々のシナプスの可塑性学習によって増強した一部のシナプスは睡眠や断眠の影響を受けずに保護された。その他のシナプス (学習により増強しなかったシナプス) が睡眠時に減弱した。
今後の研究
私たちは、平均ではなく個々のシナプスレベルで、その可塑性を捉え、長期記憶の脳表現とその変容の解明を目指しています。光遺伝学的な実験技術の発展により、特定の細胞やシナプスにおける動態の解明が可能となってきています。従来研究では、実験系をシンプルにするために、単一の学習課題の前後における睡眠時の脳動態を捉えてきましたが、今後は複数記憶の脳表現の相互作用についても取り組んでいきたいと考えています。今後の研究を通じて、「記憶を混同して覚えていることがあるのは何故か?」「眠ると頭がスッキリするのは何故か?」「沢山勉強しても脳はパンクしないのか?」といった疑問に答えることを目標としています。
用語解説
1.シナプス可塑性
神経細胞間の情報伝達の場となるシナプスの前部または後部の再構成によるシナプス伝達効率の変化。長期シナプス可塑性は学習・記憶のシナプスレベルの素過程として考えられている。
2.AMPA受容体
グルタミン酸作動性シナプスにおいて、速い興奮性シナプス伝達の大部分を担う。GluA1、GluA2、GluA3、GluA4のサブユニットからなる4量体として形成されている。
3.二光子顕微鏡
一般的な蛍光顕微鏡より波長が長い赤外線を励起光として用いるため、組織深部の観察に適している。生体動物の脳表から数100 マイクロメートル程度の深さまで観察することができる。
4.子宮内電気穿孔法
子宮の外からDNA等の核酸を胎仔に注入し、電気パルスで細胞膜の透過性を上げて神経細胞等に遺伝子発現を誘導する方法。
5.錐体細胞
大脳皮質や海馬などに存在する主要な興奮性の神経細胞。
6.樹状突起スパイン
興奮性シナプスの入力を受ける樹状突起におけるトゲ状の隆起。
7.コンプレックスホイール試験
加速回転するホイールの上でマウスを走行させる運動学習試験。マウスが縦走するホイール面に、マウスが前肢と後肢で掴むための金属棒が不均等な縦間隔で配置されている。